Summary:
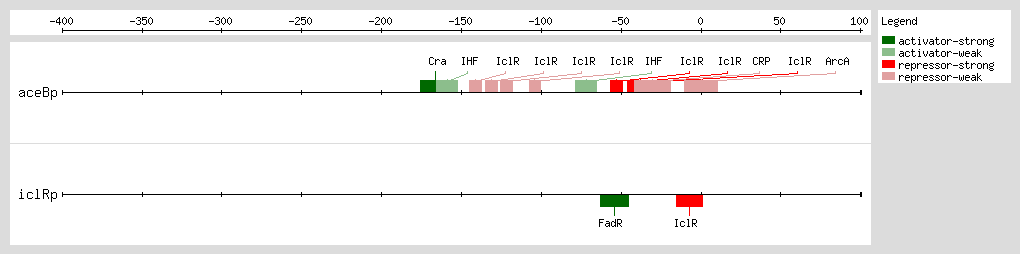
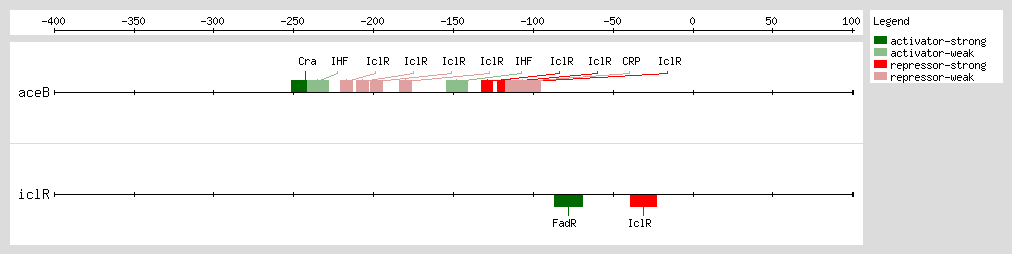
The transcription factor IclR, for "Isocitrate lyase Regulator," is negatively autoregulated [5]and it regulates the expression of the glyoxylate bypass operon [4, 6, 7, 8, 9, 10] Transcription of this operon is induced when E. coli is grown during acetate accumulation in the exponential phase. Glyoxylate and pyruvate have been identified as effectors of IclR and show antagonistic effects. While glyoxylate favors the inactive dimeric state of IclR, pyruvate increases the binding of IclR to the aceBp promoter by stabilizing the active tetrameric form of the protein [11]
On other hand, the genes of the aceBAK operon are expressed to varied degrees due to two facts: first, they are differentially regulated at the translational level, and second, there is a putative premature transcriptional termination in the region preceding the aceK gene [12]
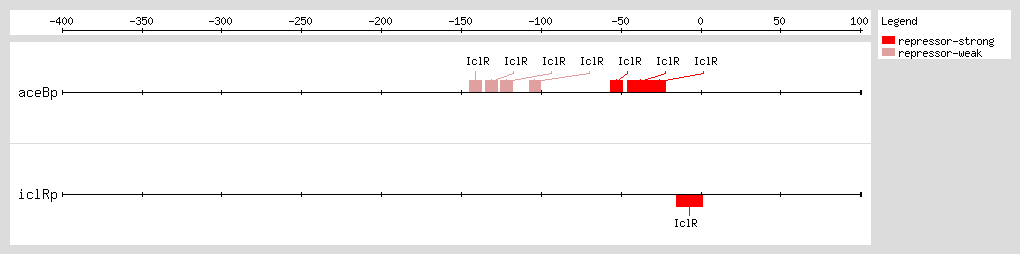
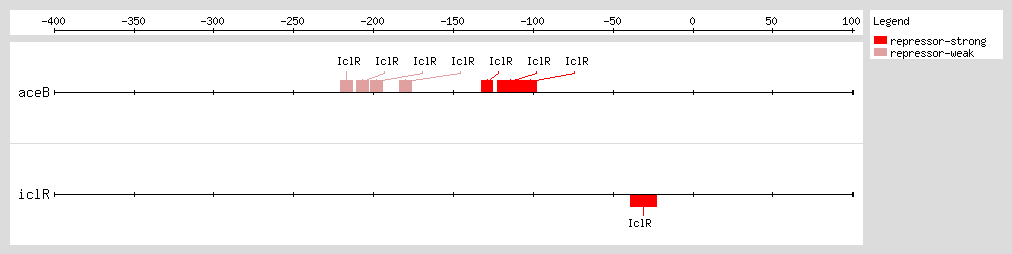
IclR represses aceBAK transcription through two mechanisms [1] (1) binding to the proximal site, overlapping the -35 promoter box, and preventing RNA polymerase binding [2, 4, 13] and (2) binding to the distal site after the RNA polymerase has bound to the promoter and formed the open complex, avoiding the polymerase escape of the promoter through its interaction with the α-subunits.
Perhaps the IclR binding at both sites forms an intermolecular bridge that leads to a DNA loop structure and thus enhances the
aceBAK repression
[1]
This regulator belongs to the IclR family of repressors. IclR is composed of two domains: the amino-terminal domain, which contains the DNA-binding region, and the carboxy-terminal domain, which is responsible for inducer binding
[11, 13, 14, 15, 16]
The crystal structure of the C-terminal domain of IclR (2.3Å) has been solved
[11]
An
iclR mutation affects the carbon flow in the metabolism
[17, 18]Under glucose-abundant conditions, a double mutant of IclR and ArcA causes an increase in biomass yield (47%) and reduction of acetate (70%) and CO
2 (16%) production. Under glucose-limited conditions, this double mutant exhibits an increase of biomass of only 13%
[19].
L-threonine production increases when the
fadR,
fabR, and
iclR regulators are deleted, suggesting that coupling fatty acid degradation and the L-threonine biosynthesis pathway via the glyoxylate shunt could efficiently increase L-threonine production
[20]
Deletion of
ilcR partially restores the growth caused by a |FRAME: EG20173| mutant under glucose depletion
[21].
Review:
[22].