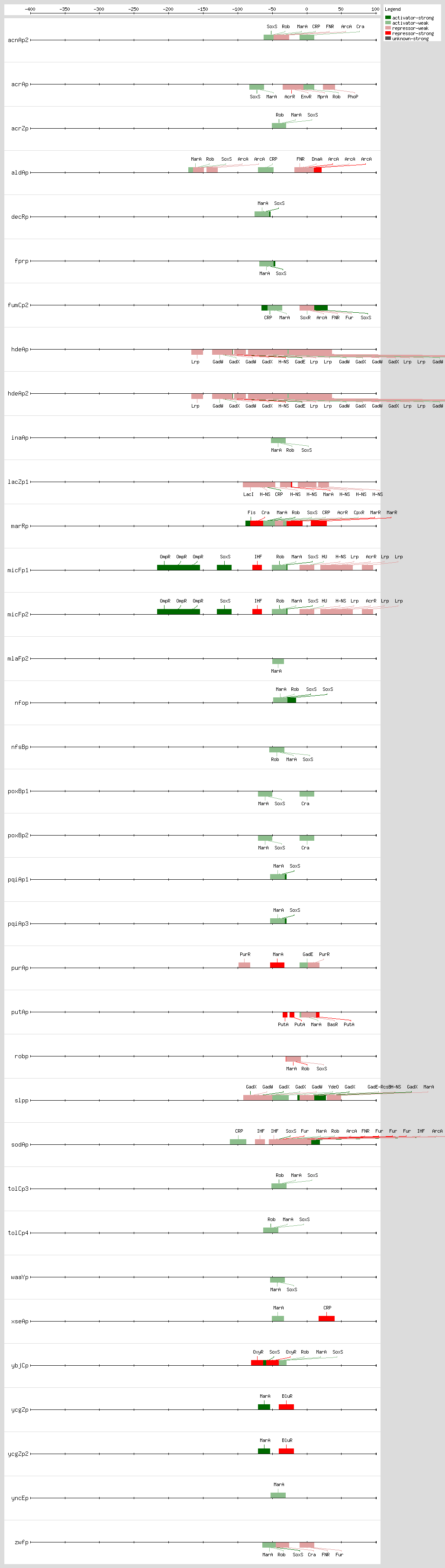
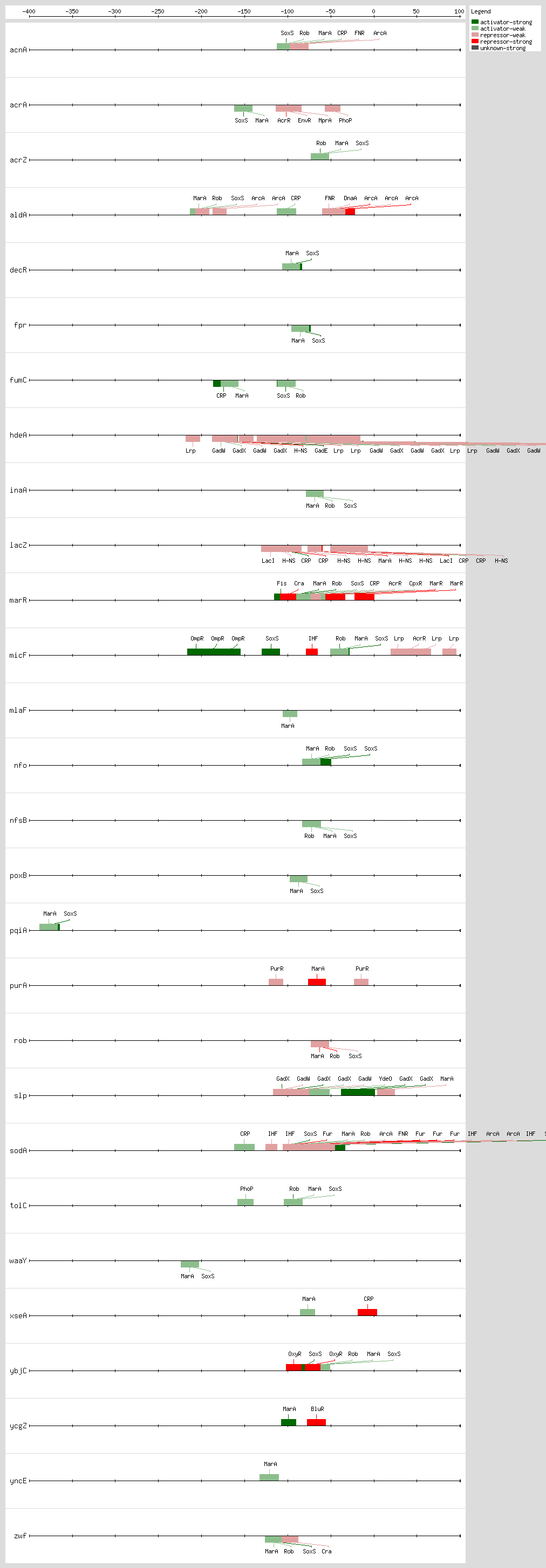
They activate a common set of genes, but the expression and activity of each one of these proteins are induced by different signals: the activity of Rob is increased with dipyridyl, bile salts, or decanoate
[31, 32], and the transcription of MarA and SoxS is increased by the aromatic weak acid salicylate
[33, 34] and oxidative stress
[35], respectively.
Many genes are regulated by all three proteins; however, some genes are regulated only by one of them. The differential regulation of these genes might be caused by the degeneracy of their DNA-binding sites
[36].
These three monomer proteins bind to the same DNA site, a degenerate 19-bp sequence known as the sox-mar-rob box, which has to be in a specific orientation and distance relative to the -35 and -10 boxes of the promoter
[3, 37]. In class I promoters, the activators bind upstream of the -35 box and are generally oriented in a backwards direction, while in class II promoters the proteins overlap the -35 promoter hexamer and generally are oriented in the forward direction
[3, 37]. In a subset of the class I promoters the sox-mar-rob box is separated by ~30 bp from the -10 hexamer but can be functional in either orientation
[3, 37].
For MarA it was shown that the extent of activation at different promoters is only poorly correlated with the strength of MarA binding to the specific mar box
[38]. By using a computational model of transcriptional activation it was proposed that at the class I promoter of
mar MarA increases the binding of RNA polymerase but not the occupancy. At the class II promoters of
sodA and
micF MarA can even decrease both RNA polymerase affinity and occupancy at the promoter. The model predicts that MarA increases the rate of transcription initiation while decreasing the overall presence of the transcription machinery at the promoter
[39].
The sox-mar-rob box contains an "invariant A" at position 1, two recognition elements, the RE1 at postion 4-7 and RE2 at position 15-18, and a 7-bp A/T-rich spacer separating these elements
[13, 40, 41]. There are more than 10,000 such binding sites per genome
[42]. However, the majority of these sites are not functional because they are not in the proper orientation or distance relative to the promoter
[33]. Based on these findings, it was proposed that these proteins activate the transcription by a mechanism named "DNA scanning" or "prerecruitment," which consists of the formation of the complex RNA polymerase transcriptional regulator in the absence of DNA, and then this complex scans the DNA to bind to appropriate sites
[33, 42].
These three proteins belong to the AraC/XylS family of transcriptional regulators
[43], and as with other members of this family they have two helix-turn-helix (HTH) motifs for DNA binding, one of them, located in the N-terminal region, interacts with the element RE1 of the mar box, and the HTH located in the C-terminal region interacts with the element RE2
[30, 40, 44]. In the case of Rob, it appears that only one of two HTH motifs makes base-specific contact with DNA; this was observed for the micF promoter
[13]. In order to decipher molecular details of the free and DNA-bound forms of Rob and MarA, some simulation analysis using native and mutant DNA binding sites and proteins has been performed
[45].
Cells carrying a mutation in
marA at a glutamic acid residue (E89A) were able to express higher resistance to superoxides than those harboring wild-type
marA [2]. Thus, MarA E89A acts more like SoxS than MarA in exhibiting greater activation and binding at the class I promoter
[2]. The MarA hydrophobic core distant from their binding site is affected by single-site substitutions, altering its extent and specificity of DNA binding
[46].
marA is the second gene of the
marRAB operon, which encodes an autorepressor (MarR) and an autoactivator (MarA)
[25]. MarR is inactivated by salicylate, and then the operon is induced. Upon removal of the inducer (salicylate), MarA is degraded by the Lon protease
[47], and the binding of MarA with DNA or RNA polymerase protects it from degradation
[48]. The transcription level of
marA was increased in the
Ion mutant and in the presence of sodium salicyclate (NaSal)
[49].
The
mar response system presents a unique antagonistic architecture, harboring weak positive feedback through MarA and a strong negative effect through MarR
[50].
The adaptability and plasticity of a regulatory network were determined by using an
in vivo system that was developed to identify variants of helix-turn-helix transcription for the MarA transcriptional regulator
[51].
Based on single-cell experiments and computational modeling, it was shown that MarA has two qualitative classes of promoters: 1) the amplifying promoters, which generate diversity and have a high channel capacity at low levels of MarA, and 2) the filtering promoters, which have low noise and only transmit information when the MarA level is high
[52].
Based on RT-qPCR, it was shown that stress resistance genes (
rpoS,
marA,
ygfA, and
relE) and antibiotic resistance genes (ARGs), especially efflux genes, are upregulated in the viable but nonculturable (VBNC) state induced in pathogenic
Escherichia coli strains by low-level chlorination exposure
[53].
Based on CRISPRi and by engineering MarA, the active degradation of MarA was analyzed. MarAs degradation impacted both the rate of coordination and the maximum coordination that downstream genes can achieve
[54].
The effects of a mutation that inactivated MarR, caused a decrease in growth fitness, and enhanced the resistance to ciprofloxacin was reverted by mutations in
marA or
lon genes, as the growth fitness was enhanced and the antibiotic resistance was decreased
[55].
The expression of
marA is increased in the presence of styrene
[56].
Reviews:
[27, 35, 57]