|
|
| Synonyms: NagC, NagC-N-acetyl-D-glucosamine 6-phosphate |
|
Summary:
The NagC, "N-acetylglucosamine," transcriptional dual regulator participates in regulating the phosphotransferase system (PTS) [18] Its function is to coordinate the biosynthesis of the amino sugars, D-glucosamine (GlcN) and N-acetylglucosamine (GlcNAc) with their catabolism [1, 2, 12, 19]. The specific inducer for NagC is GlcNAc-6-P, the product of GlcNAc transport by the PTS [1, 19] NagC is displaced from its DNA targets by interacting with GlcNAc-6-P [1] Mutation in the first two amino acids of the recognition helix of the DNA-binding motif causes GlcNAc6P to act with NagC as a corepressor instead of as an inducer [15]. Proline at the first position of this helix seems to contribute to the distinction between the NagC binding sites and suboptimal sites [15]. Based on the structure of |FRAME: PD01896|, models for the three-dimensional structure of NagC and for the binding of GlcNAc-6-P were developed [18] The Nag regulon consists of two divergent operons, nagE and nagBACD. Read more > |
| Transcription factor | ||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TF conformation(s): |
|
|||||||||||||||||||||
| Evolutionary Family: | MarR | |||||||||||||||||||||
| TFBs length: | 23 | |||||||||||||||||||||
| TFBs symmetry: | inverted-repeat | |||||||||||||||||||||
| Sensing class: | External sensing using transported metabolites | |||||||||||||||||||||
| Connectivity class: | Local Regulator | |||||||||||||||||||||
| Gene name: | nagC | |||||||||||||||||||||
| Genome position: | 700374-701594 | |||||||||||||||||||||
| Length: | 1221 bp / 406 aa | |||||||||||||||||||||
| Operon name: | nagBAC-umpH | |||||||||||||||||||||
| TU(s) encoding the TF: |
|
| Regulon |
|
||||||
|---|---|---|---|---|---|---|---|
| Regulated gene(s) | chbA, chbB, chbC, chbF, chbG, chbR, chiP, chiQ, chiZ, creA, creB, creC, creD, crr, dinI, feoA, feoB, feoC, fimB, galP, glmS, glmU, manX, manY, manZ, nagA, nagB, nagC, nagE, nanC, nanM, nanS, ptsH, ptsI, umpH, ydeM, ydeN, ydeP | ||||||
| Multifun term(s) of regulated gene(s) |
MultiFun Term (List of genes associated to the multifun term)
carbon compounds (14)
membrane (9)
Phosphotransferase Systems (PEP-dependent PTS) (8)
amino sugar conversions (5)
operon (4)
Read more >
|
||||||
| Regulated operon(s) | chbBCARFG, chiZPQ, creABCD, dinI, feoABC, fimB, galP, glmUS, manXYZ, nagBAC-umpH, nagE, nanCMS, ptsHI-crr, ydeNM, ydeP | ||||||
| First gene in the operon(s) | chbB, chiZ, creA, dinI, feoA, fimB, galP, glmU, manX, nagB, nagE, nanC, ptsH, ydeN, ydeP | ||||||
| Simple and complex regulons | AraC,GadX,NagC BasR,DksA,DksA-ppGpp,H-NS,IHF,NagC,NanR,ppGpp CRP,ChbR,NagC CRP,Cra,Mlc,NagC CRP,GalR,GalS,NagC Read more > | ||||||
| Simple and complex regulatory phrases | Regulatory phrase (List of promoters regulated by the phrase) | ||||||
| Transcription factor regulation |
|
|---|
|
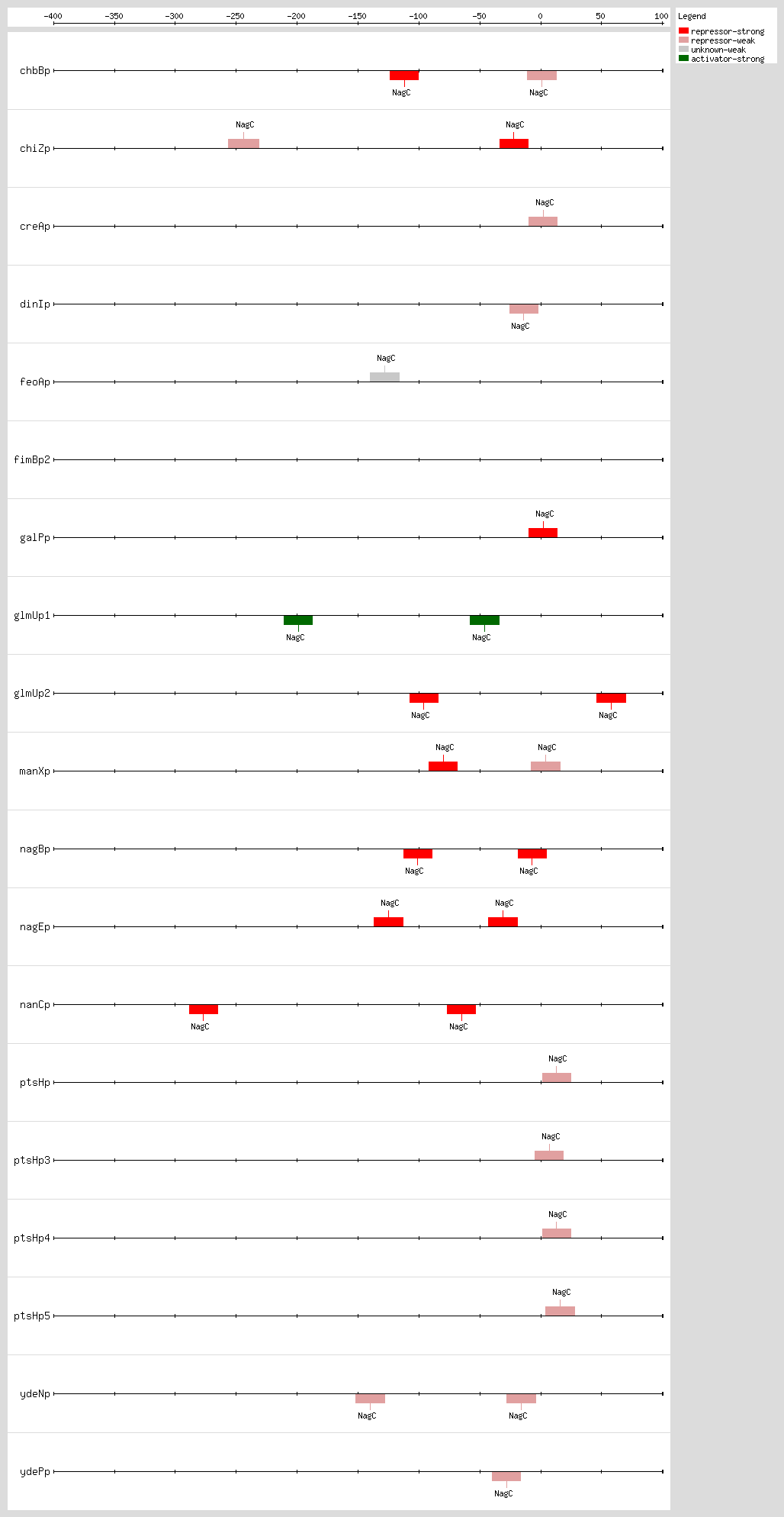
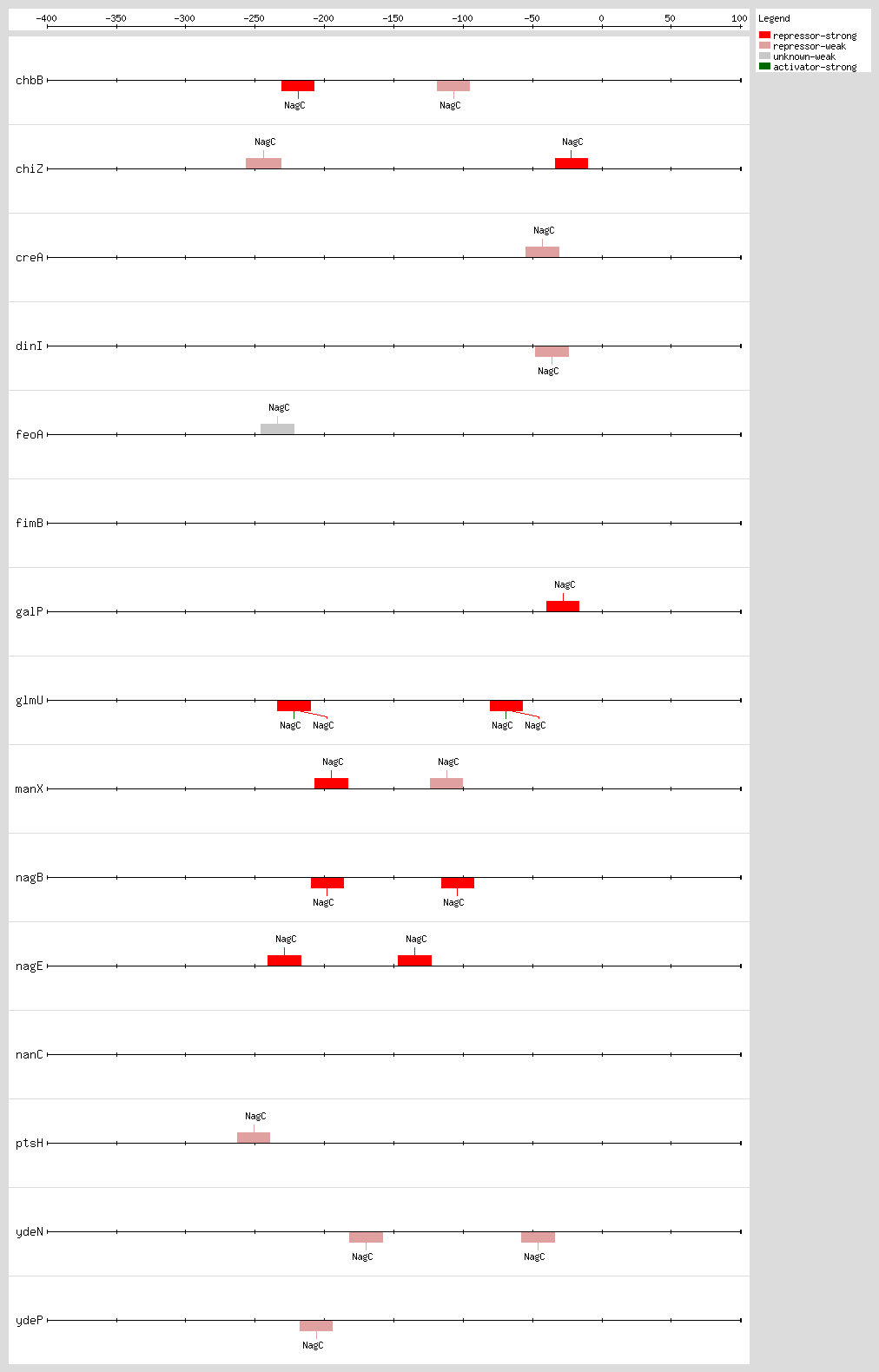
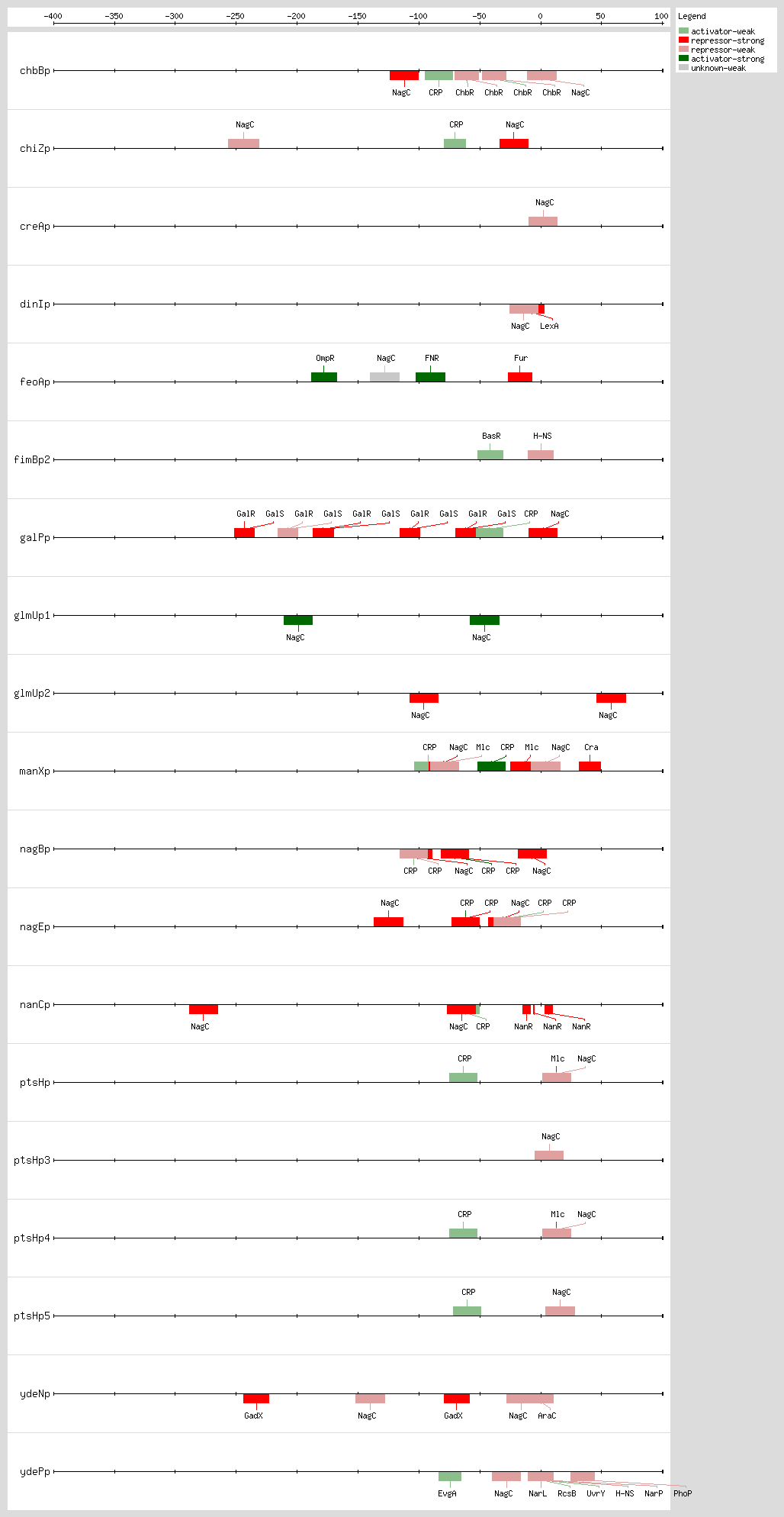
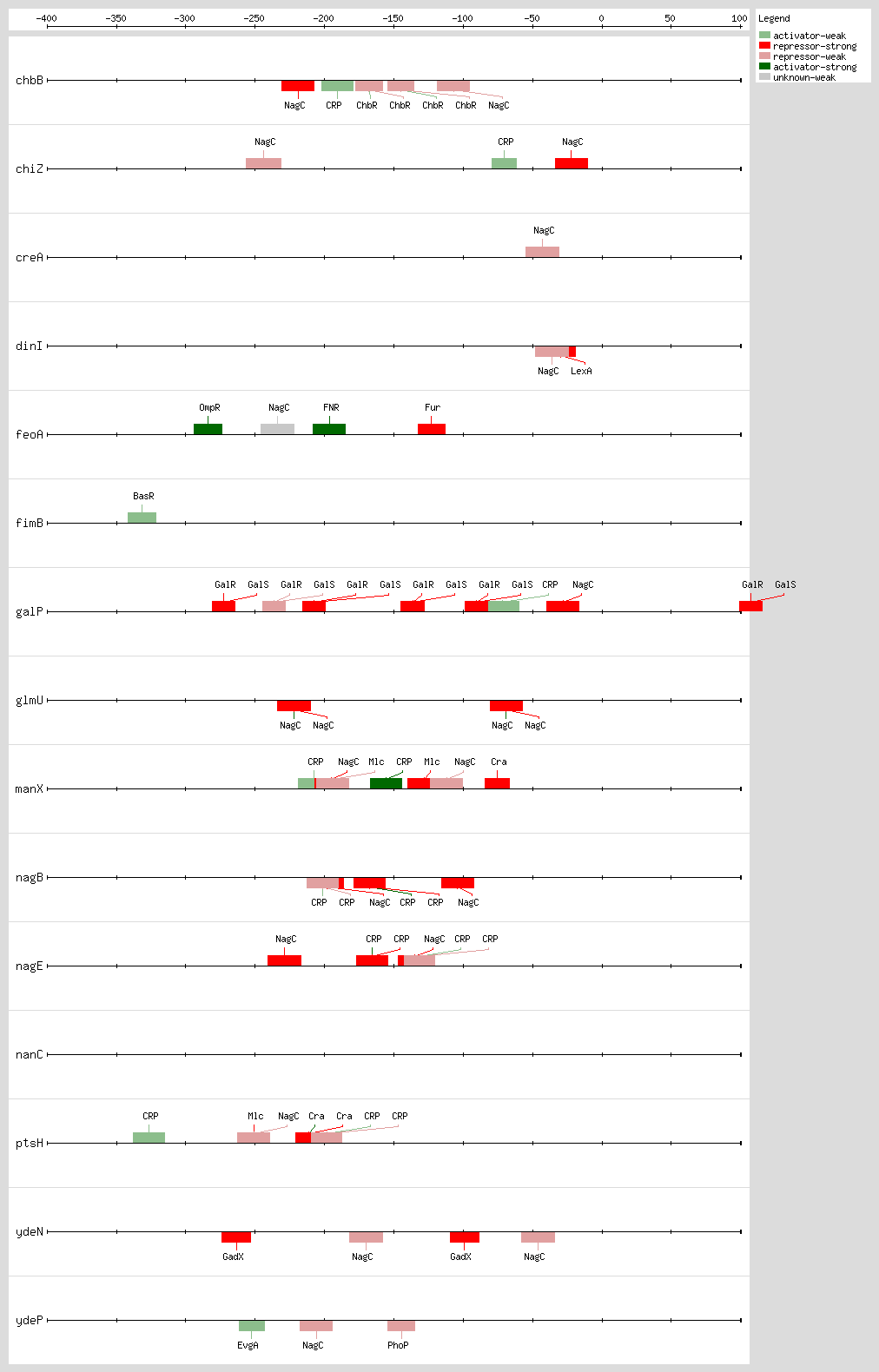
Transcription factor binding sites (TFBSs) arrangements
|
|---|
|
High-throughput Transcription factor binding sites (TFBSs)
|
|---|
| Functional conformation | Function | Object name | Object type | Distance to first Gene | Sequence | LeftPos | RightPos | Center Position | Growth Condition | Evidence | Confidence level (C: Confirmed, S: Strong, W: Weak) | References | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NagC | unknown | eutSPQTDM | Transcription-Unit | nd | nd | nd | nd | nd | [COMP-AINF-SIMILAR-TO-CONSENSUS] | nd | nd |
| Alignment and PSSM for NagC TFBSs |
|---|
| Aligned TFBS of NagC |
|---|
| Sequence | |
|---|---|
| AAATAATTCGCGAAGATAATTAAGA | |
| ACATATTTCATGATGAGAATTATGC | |
| AGTAATTTTACGAAGAGAAATAAGT | |
| CTATTTTTCGCGACGCGAATTATCA | |
| AATTAATTCGCGAAGAAAAGCAGAA | |
| ATTAATTTCGCGCATCAAAATAATT | |
| ATTTAATTCGTATCGCAAATTAAAC | |
| TCTTAATTCACAATAAAAAATAACC | |
| AATTAGTTCGTCACGGTAAATAACA | |
| AGAAATTTCGAAAGGTAAAATATCC | |
| ATATAATTCGAATAGTGAATTACCC | |
| ACATATTTTGTGACACGAATTGCAA | |
| ACTTATTTTATCATTCAAAAAATCA | |
| AGTAATTTCTCGCCGATAAACAACT | |
| ATATTTTTCAATATAAAAAATAAGA | |
| TCTTATTTCGATCAATGAATAAACA | |
| GGTTATTTTACCTGTATAAATAACC | |
| GTCCATTTCACGATGAAAAAAATGT | |
| ACTTTTTTTGTGGCTGAAAGGAGTA | |
| AAATAAATCGCGAAACGCAGGGGTT |
| Position weight matrix (PWM). NagC matrix-quality result |
|---|
A 15 4 8 4 17 6 1 0 0 7 4 4 13 7 5 9 9 19 20 10 3 18 10 3 9 C 1 6 1 1 0 0 0 0 15 0 10 3 3 6 0 6 0 1 0 0 2 0 2 9 6 G 2 4 0 0 0 1 0 0 0 12 0 11 1 2 11 2 7 0 0 3 2 2 3 5 0 T 2 6 11 15 3 13 19 20 5 1 6 2 3 5 4 3 4 0 0 7 13 0 5 3 5 |
| Consensus |
|---|
; consensus.strict acttAtTTCGcgacgagAAatAaca ; consensus.strict.rc TGTTATTTCTCGTCGCGAAATAAGT ; consensus.IUPAC aywtAwTTCRygamgmrAAwtAasm ; consensus.IUPAC.rc KSTTAWTTYKCKTCRYGAAWTAWRT ; consensus.regexp a[ct][at]tA[at]TTC[AG][ct]ga[ac]g[ac][ag]AA[at]tAa[cg][ac] ; consensus.regexp.rc [GT][CG]TTA[AT]TT[CT][GT]C[GT]TC[AG][CT]GAA[AT]TA[AT][AG]T |
| PWM logo |
|---|
|
|
|
| Evolutionary conservation of regulatory elements |
|---|
| TF-target gene evolutionary conservation |
|
| Promoter-target gene evolutionary conservation |
|
| Reference(s) |
|
|---|---|