Subsequent transfer of the phosphate group to PhoB results in activation of PhoB
[27] When phosphate is in excess, autophosphorylation of PhoR is inhibited and PhoB-P is dephosphorylated. Autoregulation of PhoB/PhoR expression serves as a mechanism for cells to achieve respective optimal densities and fitness under these conditions
[28] This negative regulation requires in addition to PhoR an intact Pst system and PhoU
[29] In the absence of PhoR, cross-regulation of PhoB by CreC or acetyl phosphate results in phosphorylation of PhoB in response to carbon sources
[30, 31] A heterodimer formed between a phosphorylated monomer and an unphosphorylated monomer of PhoB plays an important role in PhoB autophosphorylation kinetics
[32]
PhoB consists of two functional domains, the N-terminal receiver domain, which is phosphorylated, and the C-terminal output domain, which binds to DNA and interacts with the σ
70 subunit of RNA polymerase to activate transcription
[33, 34] The output domain belongs to the winged helix-turn-helix family of transcription factors
[35] Its activity is silenced by the receiver domain, and phosphorylation relieves inhibition
[36] The 3D structure of the receiver domain
[37]and of two constitutively active mutants of this domain
[38]and the 3D structure of the output domain have been solved
[39] By using motif binding geometries (MBG), a structural model of the PhoB dimer in the transcription initiation complex PhoB/RNAP/σ factor/DNA has been developed
[40]
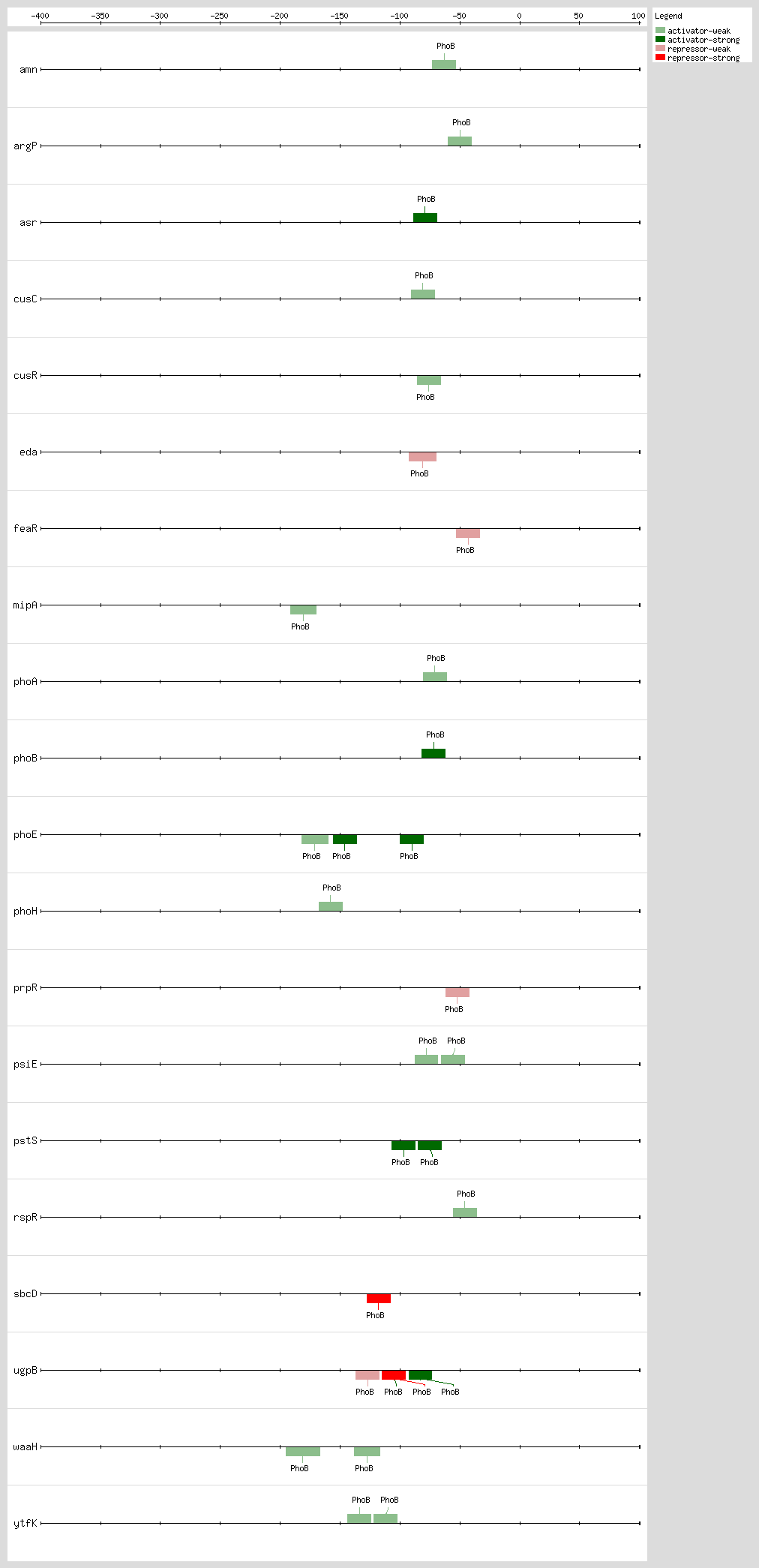
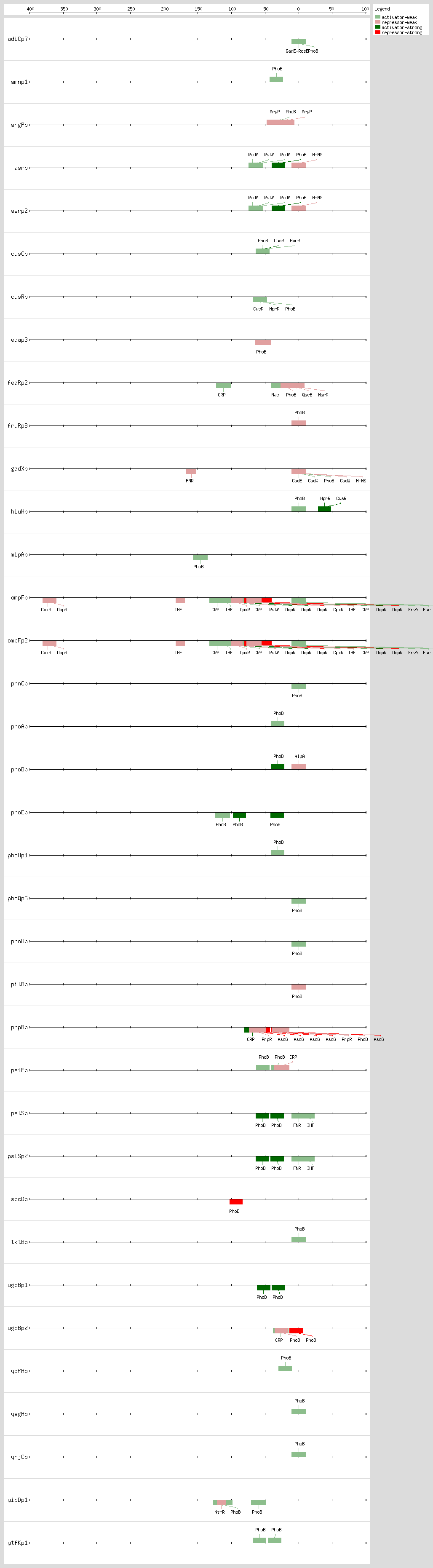
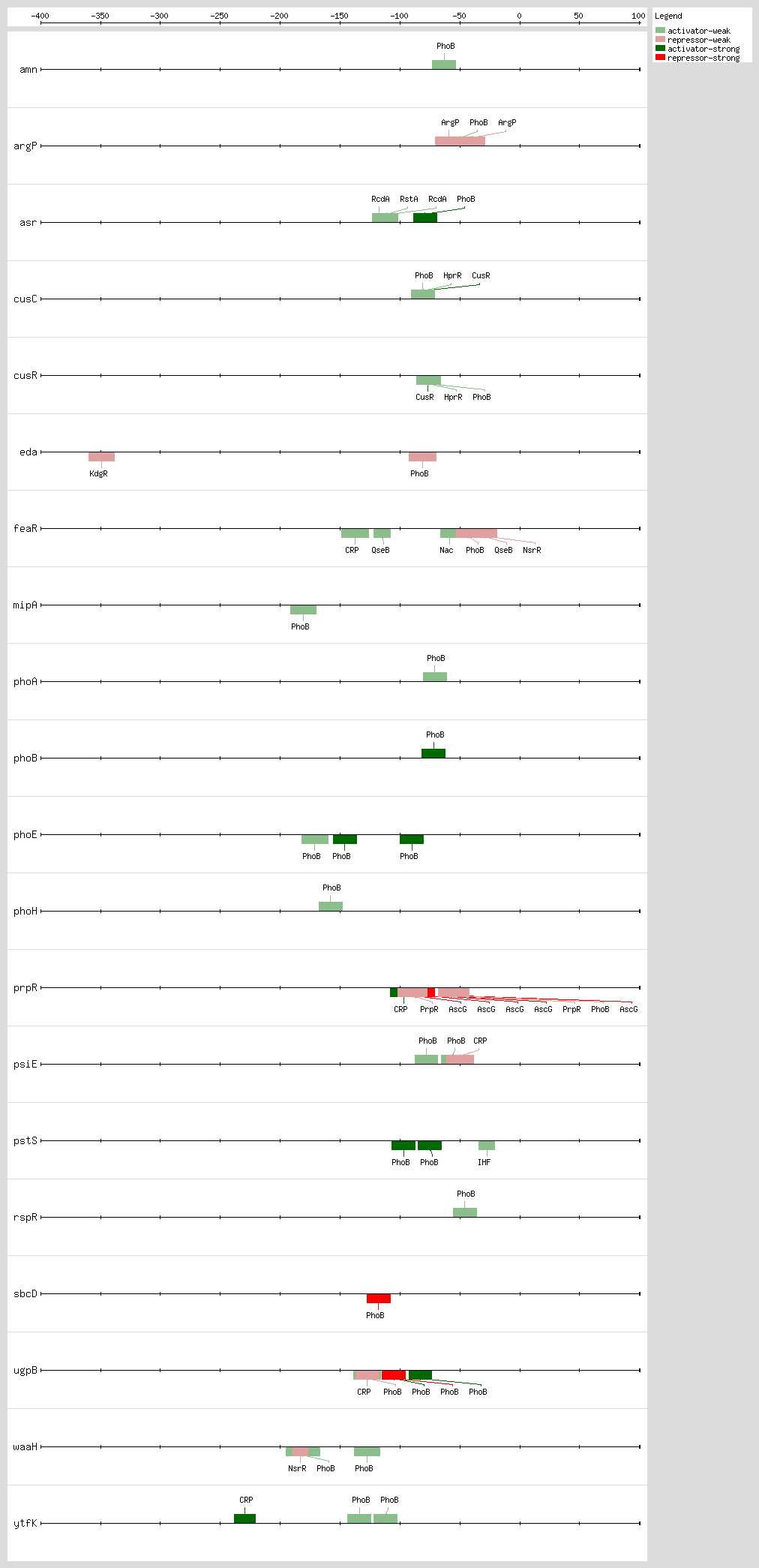
PhoB binds to the Pho box, the structure of a PhoB-DNA complex, revealing that sequence changes within the 4-bp region can alter the binding affinity without changing the major groove-contacting TGTCA tract that is important for binding specificity. Sequences are less conserved within this 4-bp region than in the rest of the 18-bp region
[41]. The 18-bp region has been described as two direct 11-bp repeats consisting of successive 7-bp direct repeats followed by an A/T-rich region of 4 bp, situated 10 bp upstream of the -10 region
[10, 18, 42] The
pho promoters contain functional -10 sequences but lack the consensus -35 sequence
[17]
The 3D structure of the PhoB effector domain in complex with its target DNA sequence reveals a novel tandem arrangement in which several monomers bind head to tail to successive 11-bp direct repeat sequences, coating one face of the double helix
[42, 43]
Based on DNA microarray analysis, it has been shown that PhoBR and PhoB are responsible for upregulation of phosphonate and glycerol phosphate metabolism and the high-affinity phosphate transport system, respectively, thus showing the complex regulation by the PhoR-PhoB two-component regulatory system
[44] The expression timing of PhoB regulation and the temporal hierarchy are related, as the genes involved in direct P
i uptake contain high-affinity sites and are transcribed earlier than genes involved in phosphorus scavenging
[41]. Although PhoB is activated under P
i limitation, it can also be activated under nonlimiting P
i conditions
[45]
PhoB is involved in the inhibition of biofilm formation that is triggered by fluctuations in poly(P) levels during the stationary phase
[45]
Based on results of a transcription reporter assay and
in vivo phosphorylation analysis, the shutoff response for the PhoR-PhoB system was determined
[46]. These comprehensive analyses revealed the impact of phosphatase activity on shutoff kinetics. The ratio of the phosphatase rate to the nonspecific phosphorylation rate correlated with TCS expression levels and the ratio of activity of the RR (PhoB response regulator) to that of the HK (histidine kinase), which may contribute to the great diversity of enzyme levels and the activities observed in different TCSs
[46].
The memory of the PhoB-PhoR two-component system (TCS) was used to study how cells cope with the recurrence of P
i starvation via a TCS. Memory prior P
i starvation can exert distinct effects through two regulatory pathways, TCS signaling and the stress response
[47] The crystal structures for inactivated and BeF
3-activated PhoB
N (N-terminal receiver domain) were solved by using X-ray at 2.4 and 1.9 A resolution, respectively
[48] The backbone assignment of a PhoB
NF20D mutant has been determined using NMR spectroscopy, and this information could be useful for studying the structure and dynamics of the PhoB receiver domain and its mechanism of phosphorylation in TCSs
[48] Based on NMR spectroscopy, the apo form of the N-terminal receiver domain (PhoB
N) in solution was determined
[49] In addition, it was shown via concentration dilution experiments and backbone relaxation parameters that PhoB
N exists in equilibrium between monomer and dimer states. Finally, paramagnetic relaxation enhancement (PRE) measurements showed that the dimer of apo PhoBN forms several transient dimer interfaces in solution
[49]
Based on wild-type and recombinant cells cultivated in two different substrates (lactose and glycerol), the effect of phosphate-starved response (PSR) stimuli was investigated. Gene expression analysis showed that under PSR conditions, selected genes (
gapA, pykF, ppsA, icdA) in the glycolysis and pentose phosphate pathways (
zwf, gnd, talB, tktA) were up-regulated, whereas other genes in lactose (
lacY, lacA) and acetate (
ackA, pta) metabolism pathways were down-regulated. It was also demonstrated that Cra, CRP transcription factors, and the PhoBR two-component signal transduction system are involved in the regulation of central carbon metabolism
[50]