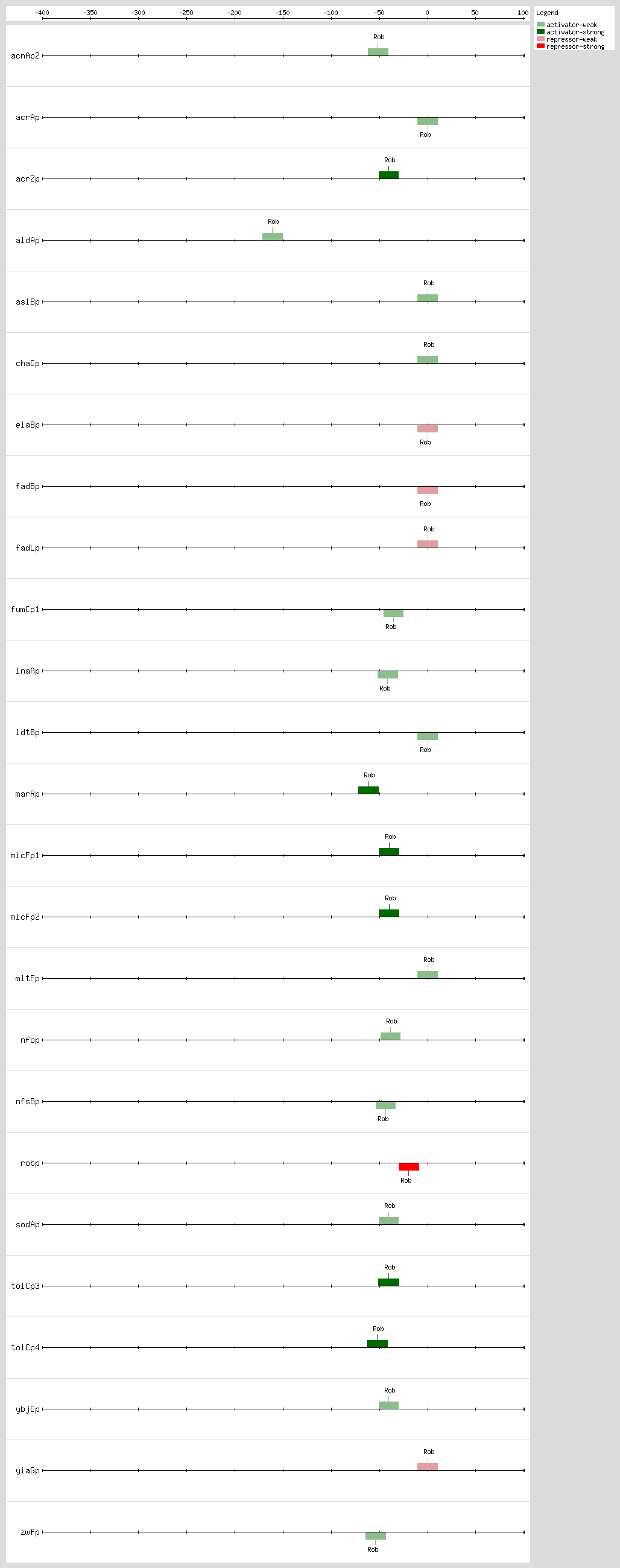
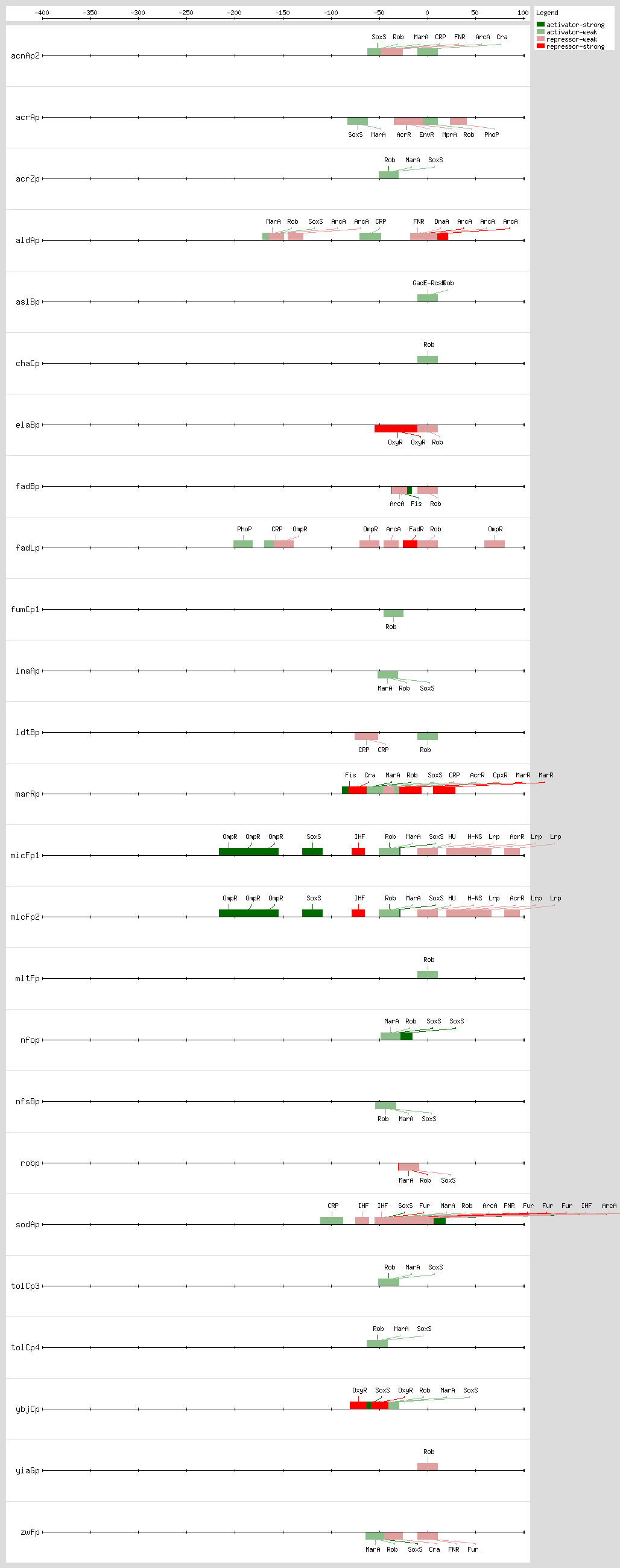
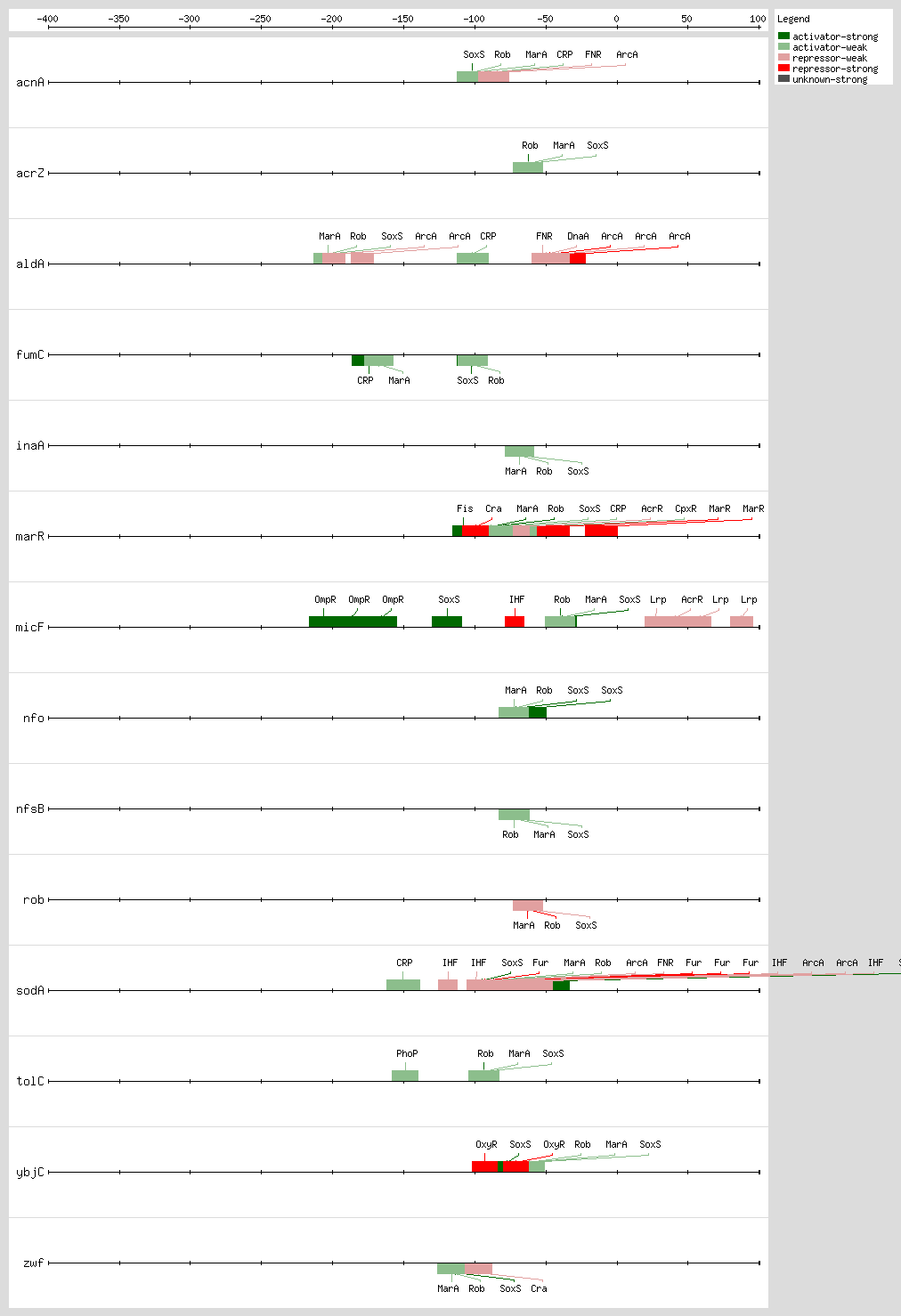
Cross talk between the
mar and
rob systems plays an important role in the response to salicylate
[33]
Many genes are regulated by all three proteins; however, some genes are regulated by only one of them. The differential regulation of these genes might be caused by the degeneracy of their DNA-binding sites
[34]
Rob, MarA, and SoxS bind as monomers to the same DNA site, a degenerate 20-bp sequence known as the Mar-Sox-Rob box, which has to be in a specific orientation and distance relative to the -35 and -10 boxes of the promoter
[9, 35] In class I promoters, the activators bind upstream of the -35 box and are generally oriented in the backward direction, while in class II promoters the proteins overlap the -35 promoter hexamer and generally are oriented in the forward direction
[8, 9, 35] As a subset of the class I promoters, the Mar-Sox-Rob box is separated by ~30 bp from the -10 hexamer but can be functional in either orientation
[9, 35]
The Mar-Sox-Rob box contains an "invariant A" at position 1, two recognition elements, the RE1 at position 4-7 and RE2 at position 15-18, and a 7-bp A/T-rich spacer separating these elements
[12, 36, 37] There are more than 10,000 such binding sites per genome
[38] However, the majority of these sites are not functional because they are not in the proper orientation or distance relative to the promoter
[39] It was proposed that these proteins bind to their target sites by a mechanism named "DNA scanning" or "prerecruitment." In prerecruitment, the protein first binds to RNA polymerase in solution, and the binary complex then scans the DNA to find its binding sites
[38, 39]
Rob consists of two domains and belongs to the AraC/XylS family
[40] The N-terminal domain is the DNA-binding domain and is homologous to MarA and SoxS and the C-terminal domain of AraC. It carries two helix-turn-helix (HTH) motifs for DNA binding. One of them, located in the N-terminal region, interacts with the element RE1 of the Mar-Sox-Rob box, and the HTH located in the C-terminal region interacts with RE2
[36, 41, 42] In the case of Rob, it appears that only one of the two HTH motifs makes base-specific contacts with DNA
[12] In order to decipher molecular details of the free and DNA-bound forms of Rob and MarA, some simulation analysis using native and mutant DNA binding sites and proteins has been performed
[43]. On the other hand, the crystal structures of Rob
[12]and MarA
[42]in complex with DNA and the solution structure of the DNA-binding domain of AraC
[44]have been solved.
The C-terminal domain of Rob is similar to a portion of the enzyme GalT
[12] It regulates the activity of Rob by a "sequestration-disposal" mechanism, that is, it mediates the sequestration of Rob into intracellular foci. Sequestered Rob is inactive. Upon addition of the inducer, Rob is released in its active form
[45] The C-terminal domain blocks in addition proteolytic degradation by Lon protease
[45]
rob appears to be transcribed abundantly at 5,000-10,000 molecules per cell
[46, 47] throughout the growth cycle.
rob expression is enhanced at the stationary phase and under glucose and phosphate starvation, and it was suggested that expression of rob is σ
S dependent
[48]
The Rob transcription factor modified
in vivo in the presence of nucleobase amino acids at position 40 exhibited enhanced DNA promoter binding as well as phenotypic changes including resistance to antibiotics
[49]
The replacement of Arg90 with purine nucleobase amino acids increases the affinity of Rob for its DNA binding site and results in moderate phenotypic changes in resistance to antibiotics and HgCl
2 [50].
The inactivation of Rob provokes changes in the expression of 285 genes and causes greater tolerance to butanol
[6].
In cells induced for chlorhexidine (CHX) resistance, expression levels of |FRAME: EG10958| and
rob are decreased
[51].
Reviews:
[32, 52, 53]