The activity of each protein is induced by different signals: the activity of Rob is increased with dipyridyl, bile salts, or decanoate
[46, 47], and the activities of MarA and SoxS are increased by the aromatic weak acid salicylate
[6, 48] and by oxidative stress
[32], respectively. SoxS was induced by tellurite (TeO
32-) in a DNA microarray analysis
[49].
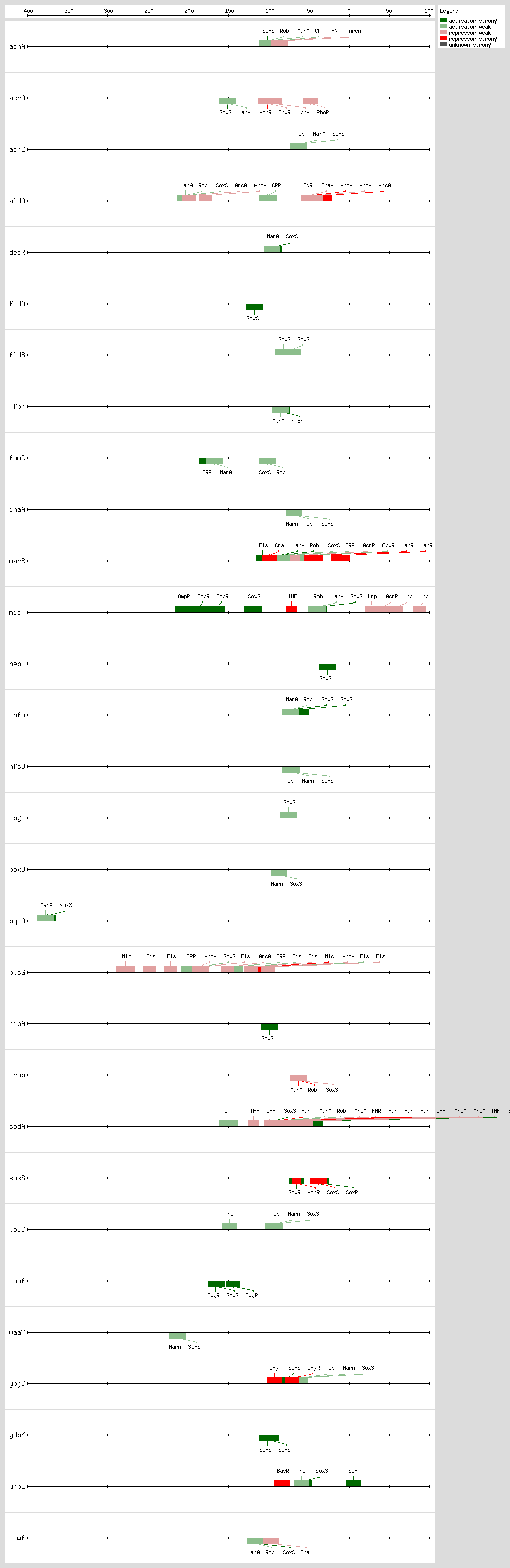
Many genes are regulated by all three proteins; however, some genes are regulated by only one of them. The differential regulation of these genes might be caused by the degeneracy of their DNA-binding sites
[50].
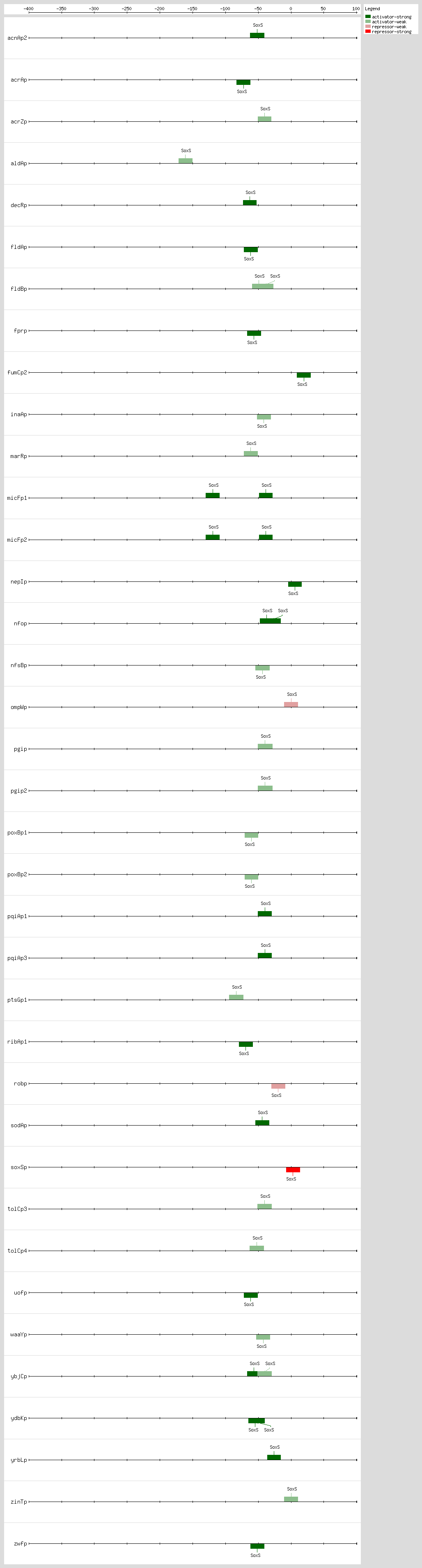
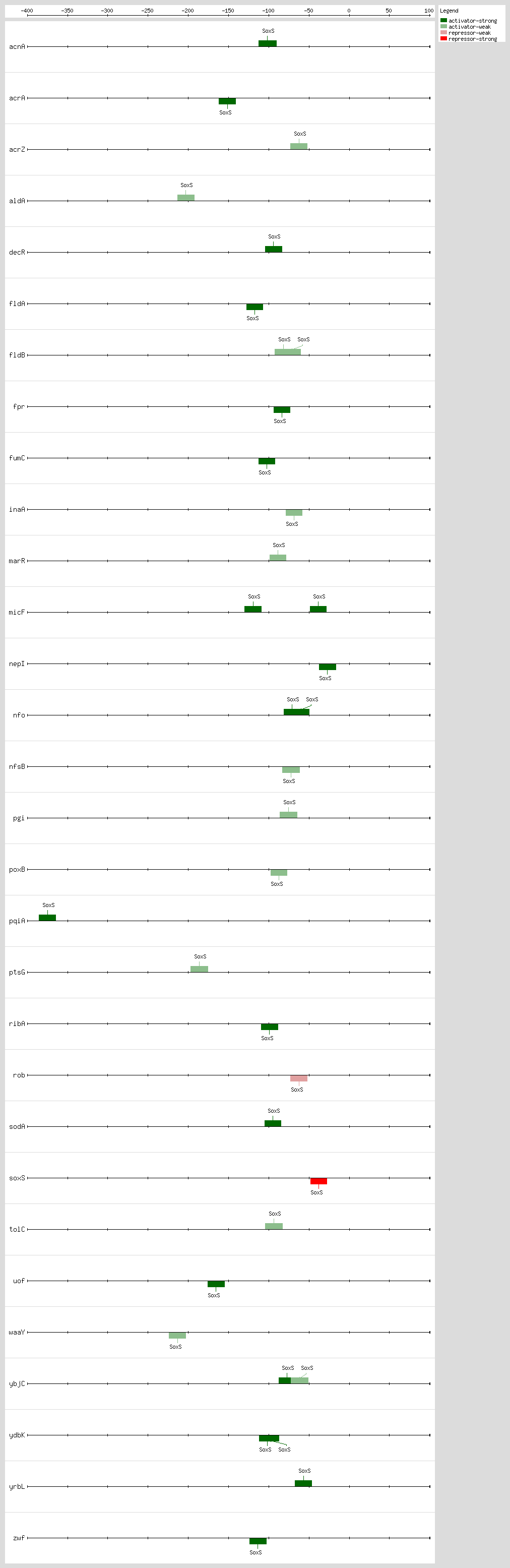
SoxS, MarA, and Rob bind as monomers to the same DNA site, a degenerate 20-bp sequence known as the Mar-Sox-Rob box, which has to be in a specific orientation and distance relative to the -35 and -10 boxes of the promoter
[4, 51]. In class I promoters, the activators bind upstream of the -35 box and are generally oriented in the backward direction, while in class II promoters the proteins overlap the -35 promoter hexamer and generally are oriented in the forward direction
[4, 51]. As a subset of the class I promoters, the Mar-Sox-Rob box is separated by ~30 bp from the -10 hexamer but can be functional in either orientation
[4, 51].
The Mar-Sox-Rob box contains an "invariant A" at position 1, two recognition elements, the RE1 at position 4-7 and RE2 at position 15-18, and a 7-bp A/T-rich spacer separating these elements
[52, 53, 54]. There are more than 10,000 such binding sites per genome
[55]. However, the majority of these sites are not functional because they are not in the proper orientation or distance relative to the promoter
[56]. It was proposed that these proteins bind to their target sites by a mechanism named "DNA scanning" or "prerecruitment." In prerecruitment, the protein first binds to RNA polymerase in solution, and the binary complex then scans the DNA to find its binding sites
[55, 56]. It has been shown that SoxS binds to the C-terminal domain of the α-subunit of RNA polymerase (α-CTD), in particular, it interacts with amino acid residues lying in or near the DNA-binding determinant of α-CTD. Apparently, SoxS functions as a co-sigma factor and redirects RNA polymerase to promoters carrying a SoxS-binding site
[57].
A new interaction that also occurs via prerecruitment between σ
70 R4 of RNA polymerase (RNAP) and SoxS has been identified
[58]. There is evidence for the existence of two types of interactions between SoxS and σ
70 R4, with one type being able to occur "off-DNA" (within a SoxS-RNAP binary complex that scans the chromosome for SoxS-dependent promoters) and with the other one occurring "on-DNA" (within a SoxS-RNAP binary complex residing at a SoxS-dependent promoter)
[58]. SoxS is now the first example of an
Escherichia coli transcriptional activator that uses a single positive control surface to make specific protein-protein contacts with two different subunits of RNAP
[58]. In class I promoters, the σ
70 R4 region binds to the -35 box and SoxS binds upstream of the hexamer, but in class II promoters SoxS binds to the -35 box, occluding the binding of σ
70 R4
[59].
SoxS, MarA, and Rob belong to the AraC/XylS family of transcriptional regulators
[60] and, like other members of this family, they have two helix-turn-helix (HTH) motifs for DNA binding: one of them, located in the N-terminal region, interacts with the element RE1 of the Mar-Sox-Rob box, and the HTH located in the C-terminal region interacts with the element RE2
[53, 61, 62].
soxS is divergently transcribed from
soxR [63]. SoxR is the transcriptional activator for
soxS and a sensor for superoxide and nitric oxide through its [2Fe-2S] center. It activates
soxS transcription when oxidized
[64, 65].
soxS is regulated only by its intracellular concentration. Upon removal of the inducer, SoxS is degraded by the Lon protease
[66]. The binding of SoxS with DNA or RNA polymerase protects it from degradation
[67].
Transcriptomic and statistical modeling analyses with Zn concentration shifts showed a transient change in the activity of the iron regulator Fur and of the oxidative stress regulator SoxS; these findings suggest a competitive binding mechanism for the physiology of Zn and Fe uptake
[68]. The expression of
soxS is not affected by iron limitation
[17].
Cadmium prevented the induction of
soxRS-controlled protective enzymes by the redox-cycling agent paraquat
[69], although this appeared not to be due to specific inactivation of the
soxRS regulon and most probably resulted from generalized suppression of protein synthesis
[69, 70].
The expression of the gene
soxS is enhanced in the presence of a copolymer based on monomethylether poly(ethylene glycol), methyl methacrylate, and 2-dimethyl(aminoethyl) methacrylate
[71].
In cells induced for chlorhexidine (CHX) resistance, expression levels of
soxS and |FRAME: EG11366| are decreased
[72].
Reviews:
[32, 73, 74, 75]